 New Products
New Products Earth-Friendly Products
Earth-Friendly Products Biotium Choice Antibodies
Biotium Choice Antibodies Special Offers
Special Offers







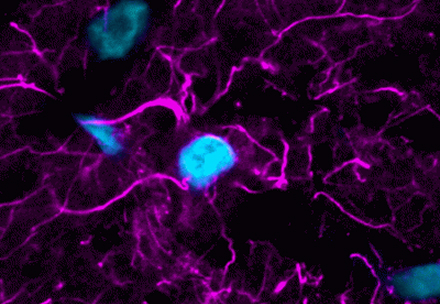
Content #1
Content #1
Content #1
A thorough understanding of biological systems requires more than identifying which molecules are present; it requires understanding how they are organized and interact within intact tissue environments. This principle underlies spatial biology, a rapidly growing field that integrates molecular profiling with spatial context to provide a systems-level view of cellular organization, function, and interaction.1-2 Unlike standard immunohistochemistry and immunofluorescence approaches, which typically examine only one or a few markers at a time, modern spatial biology combines high-dimensional molecular profiling, multiplex imaging, and computational analysis to map complex cellular interactions and tissue architecture within intact samples.1-2 These techniques generate multidimensional spatial datasets that provide deeper insight into cellular organization and function across diverse biological systems.
Bulk and single-cell omics approaches have revolutionized molecular profiling but often require tissue dissociation, disrupting the spatial organization that shapes cellular behavior.1-2 Conversely, conventional tissue staining techniques preserve tissue architecture but have historically been limited in multiplexing capacity and molecular depth. Spatial biology emerged from the convergence of imaging, sequencing, and computational technologies to overcome these limitations by enabling high-dimensional analysis of RNA, proteins, and other biomolecules directly within intact tissues.1 These approaches generate multidimensional spatial datasets that map molecular profiles and cell-cell interactions in situ, providing systems-level insight into tissue organization and function.1
Spatial context is essential for studying processes in which cellular organization plays a defining role, including:
At the same time, the complexity of these systems has driven the need for technologies capable of detecting increasing numbers of molecular targets within a single sample.
Central to many spatial biology workflows is multiplexed imaging, which enables simultaneous detection of multiple targets within intact tissues.2 While multiplex imaging has existed for decades, the increasing complexity of spatial biology applications has pushed these technologies to evolve to enable higher-plex and more information-rich analyses.2 Here, we’ll explore a selection of those techniques with an emphasis on fluorescence-based imaging techniques (Table 1).
Table 1. Comparison of Multiplexing Strategies
| Multiplex IHC | Multiplex IF (mIF) | Spectral Imaging | Cyclic IF | TSA-Enhanced Cyclic IF | |
|---|---|---|---|---|---|
| Detection modality | Enzyme- mediated chromogen deposition | Direct/indirect fluorescence | Multispectral fluorescence with unmixing | Iterative fluorescence staining | HRP-mediated tyramide deposition (covalent) |
| Multiplex capacity | ~4-8 | ~4-8 | ~6-24+ | ~20-60+ | ~20-60+ |
| Workflow Complexity | Single round | Single round | Single round | Multi-cycle (stain-image-strip) | Multi-cycle with signal retention |
| Key Advantage | Simplicity; clinical compatibility; lower barrier to entry in terms of equipment | Higher sensitivity vs IHC | Resolves partially overlapping spectra | Scalable multiplexing | High sensitivity and signal stability |
| Key Limitation | Limited plex; chromogen overlap | Spectral overlap; antibody constraints | Spectral complexity ceiling; requires costly equipment and softwear | Tissue stress; signal loss; residual signal carryover; costly automated staining and scanning equipment (if used) | Reagent stability; tissue stress; workflow complexity; residual signal carryover; costly automated staining and scanning equipment (if used) |
| Signal Stability | Permanent | Moderate (photobleaching) | Moderate | Limited across cycles | Strong (covalent deposition) |
| Ideal Application | Low-plex pathology | Moderate multiplex profiling | Expanded low-plex panels | High-dimensional phenotyping | High-plex, low-abundance targets |
Spatial omics encompasses a group of high-throughput molecular profiling techniques that measure and map the spatial distribution of RNA, proteins, and other biomolecules within intact tissues.1 Spatial transcriptomics is a branch of spatial omics that measures gene expression by detecting RNA molecules within intact tissue sections.1 In imaging-based spatial transcriptomics workflows, tissue sections are prepared and processed to preserve tissue morphology while enabling RNA accessibility. Target-specific probes hybridize to RNA transcripts within the tissue, followed by ligation and rolling circle amplification to generate localized amplification products that enhance fluorescence-based detection (Fig. 1).1-2 Histological or fluorescence-based staining may also be performed to visualize tissue morphology and support cell segmentation. Fluorescent readout probes are then sequentially hybridized and imaged across multiple imaging cycles to identify spatially resolved gene expression patterns within the tissue section. Computational decoding and image analysis reconstruct transcript localization and cellular organization within the tissue microenvironment.1-2
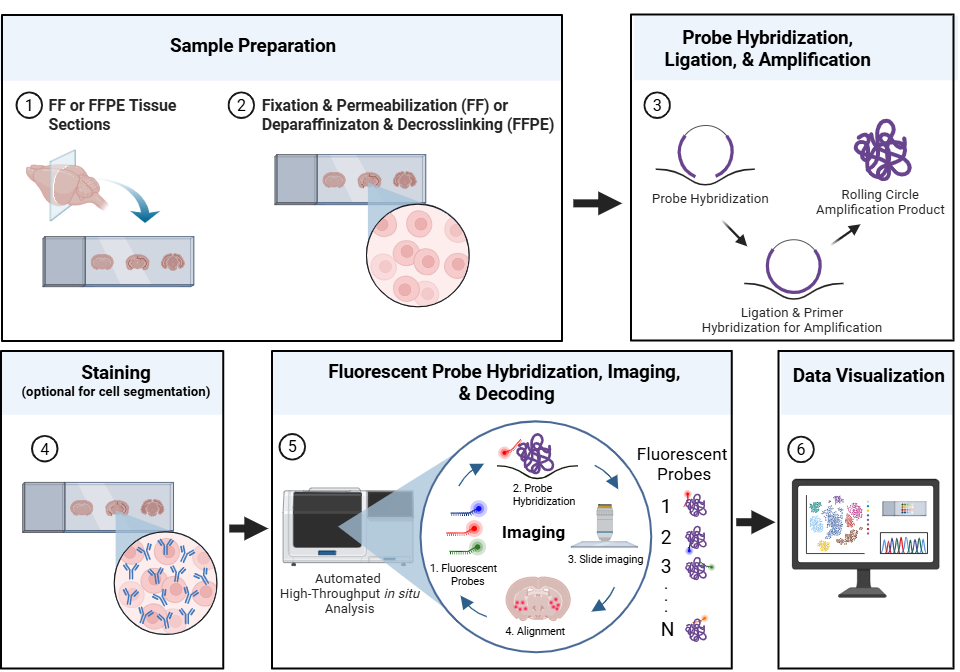
Figure 1. A representative workflow for imaging-based spatial transcriptomics. Fresh frozen (FF) or formalin-fixed paraffin-embedded (FFPE) tissue sections are prepared and processed to preserve tissue morphology and enable RNA accessibility. Target-specific probes hybridize to RNA transcripts within the intact tissue, followed by ligation and rolling circle amplification to generate localized amplification products. Histological or fluorescence-based staining may be performed to support tissue visualization and cell segmentation. Fluorescent readout probes are then sequentially hybridized and imaged to identify spatially resolved gene expression patterns within the tissue section. Computational decoding and visualization reconstruct transcript localization and cellular organization within the tissue microenvironment. Adapted from 10x Genomics spatial transcriptomics workflow overview.8 Davis, J. (2026) https://BioRender.com.
Methods such as fluorescence in situ hybridization (FISH), including highly multiplexed variants such as seqFISH, use labeled probes to localize specific transcripts, while more advanced approaches enable the simultaneous detection of hundreds to thousands of RNA species.2 These technologies provide insight into gene expression programs and cellular organization within complex tissues.2
Spatial proteomics complements transcriptomics by mapping protein distribution.1 Immunofluorescence-based methods use antibodies to detect and localize proteins at single-cell or subcellular resolution and are widely accessible on standard microscopy platforms. To expand multiplexing capacity, alternative approaches have been developed. Imaging Mass Cytometry (IMC™) from Standard BioTools uses metal-labeled antibodies to measure dozens of proteins simultaneously without spectral overlap, while DNA-barcoded antibody systems, such as Akoya Biosciences’ PhenoCycler™ (formerly CODEX®) platform, enable highly multiplexed detection through iterative imaging or sequencing readouts1 A key advantage of spatial proteomics is its ability to detect post-translational modifications, which are critical for protein function but not captured by RNA-based methods.1
Integrating spatial transcriptomics and proteomics provides a more complete, systems-level view of cellular biology. Because mRNA abundance does not always correlate with protein expression, combining these approaches allows researchers to link gene expression programs with their functional protein outputs within the same spatial context.1-2
Single-marker IHC provides target localization in tissue but often fails to capture the complexity of cellular interactions.4 Multiplex immunohistochemistry (IHC) addresses this gap by allowing for the analysis of multiple biomarkers in situ, offering a more complete view of tissue organization and disease pathology.2, 4
Multiplex IHC enables the detection of multiple protein biomarkers using distinct chromogens within a single tissue section, providing insight into protein localization and its relationship to disease state.4 This supports a more comprehensive characterization of tissue architecture, including the identification of different cell types and their spatial relationships. However, chromogenic multiplexing remains limited in plex capacity and is less scalable for high-dimensional analysis compared to fluorescence-based approaches.1, 4
Multiplex immunofluorescence (mIF) extends these capabilities by using fluorophore-labeled antibodies to detect multiple targets within the same tissue section.2 In its simplest form, mIF involves simultaneous staining with multiple antibodies, each conjugated to a spectrally distinct fluorophore.2 As with other antibody-based imaging techniques, antibody selection and validation are critical for ensuring specificity and reproducibility. Fluorophore properties, including excitation and emission spectra, must be carefully chosen to minimize spectral overlap and background signal. In multiplex workflows, these challenges become more complex because multiple antibodies and fluorophores are used simultaneously, increasing the risk of species cross-reactivity, nonspecific binding, spectral overlap, and background signal.
To address spectral overlap challenges, many high-plex imaging workflows incorporate spectral imaging and computational spectral unmixing approaches, which mathematically separate overlapping fluorescence signals and tissue autofluorescence into their individual spectral components.4 Spectral unmixing can improve signal discrimination and expand multiplexing capacity beyond the limits of conventional fluorescence microscopy. However, the finite number of spectrally distinguishable fluorophores, along with challenges related to spectral overlap and tissue autofluorescence, still places practical limits on how many targets can be simultaneously imaged within a single staining cycle.4 These limitations have driven the development of strategies such as cyclic immunofluorescence.
To overcome the spectral limitations of conventional immunofluorescence, many spatial biology workflows now incorporate cyclic or iterative imaging strategies. Rather than detecting all targets simultaneously, cyclic immunofluorescence (CycIF) uses repeated rounds of staining, imaging, and fluorophore inactivation or signal removal to expand multiplexing capacity within a single sample (Fig. 2).2-3 This approach enables the detection of far more biomarkers than would be possible using conventional fluorescence imaging alone.2-3
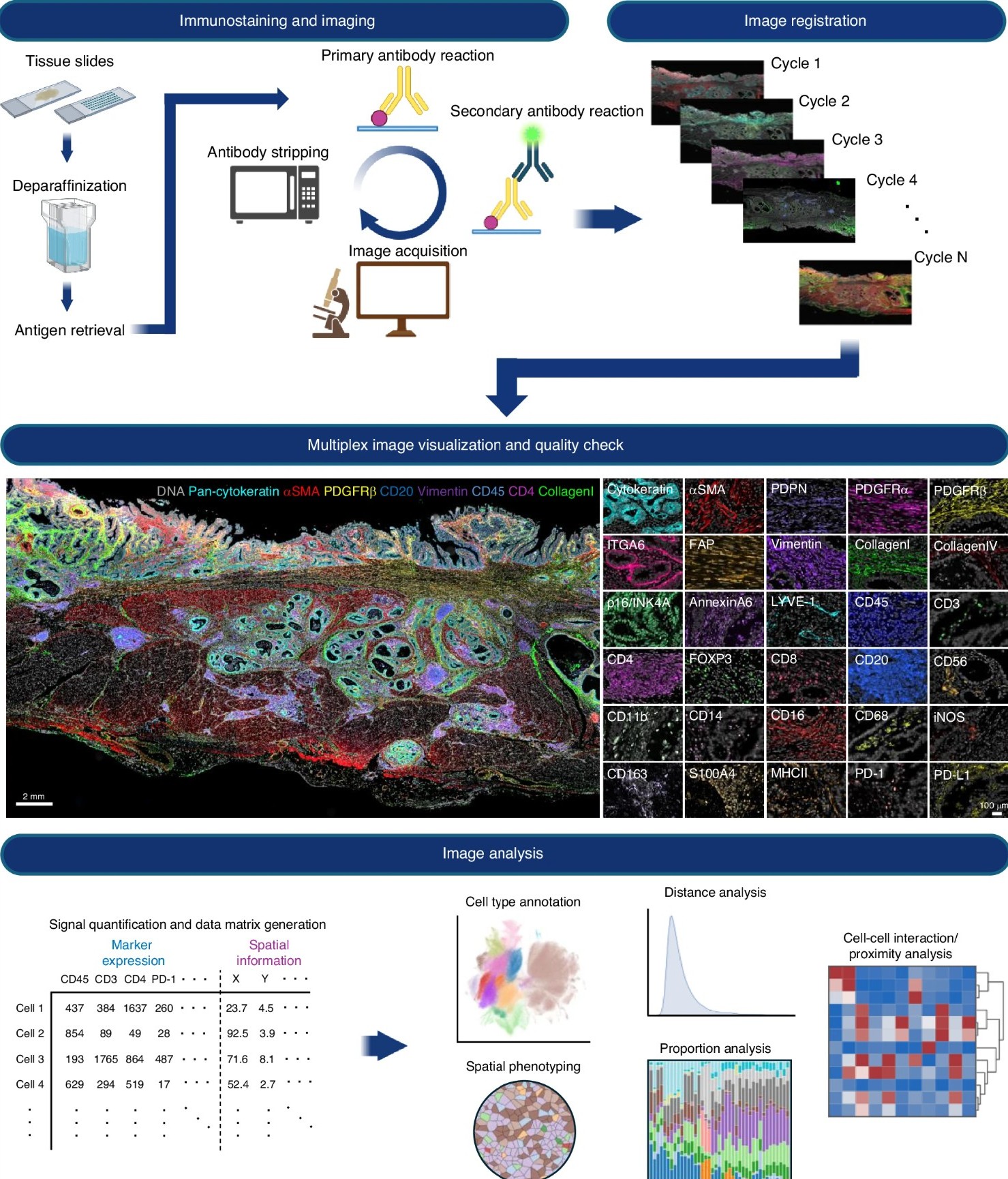
Figure 2. Current example of a CycIF multiplex imaging and analysis workflow. Reproduced from Semba T, Ishimoto T. Br J Cancer. 2024;131:1737-1747. https://doi.org/10.1038/s41416-024-02882-6. Licensed under CC BY 4.0.
In a typical CycIF workflow, a tissue section or cultured cell sample is stained with a small panel of fluorophore-conjugated antibodies and imaged using standard fluorescence microscopy.3 After imaging, the fluorescent signal is chemically inactivated or removed while preserving the underlying tissue architecture and antibody targets. Additional rounds of staining and imaging can then be performed on the same sample, allowing researchers to iteratively build high-dimensional datasets containing dozens of protein markers.3
Sequential staining approaches offer several advantages for multiplex imaging. Because only a limited number of fluorophores are used during each imaging cycle, these workflows reduce spectral overlap and simplify panel design.3 Sequential workflows can also minimize antibody cross-reactivity compared to simultaneous multiplex staining approaches, including enabling the use of multiple primary antibodies raised in the same host species across separate imaging cycles. This can improve staining flexibility and specificity in complex tissue samples. However, cyclic imaging workflows also introduce technical challenges. Repeated rounds of staining, washing, imaging, and antibody stripping or fluorophore inactivation can increase the risk of tissue degradation and sample loss over time.3 Careful optimization of fixation conditions, antibody validation, fluorophore selection, and image registration (i.e., the computational alignment of sequential images allowing for accurate overlay) is therefore critical for maintaining sample integrity and data consistency across imaging cycles.
As multiplex imaging panels continue to expand, detecting low-abundance biomarkers while maintaining strong signal specificity and spatial resolution becomes increasingly challenging. Tyramide signal amplification (TSA) is an enzyme-mediated amplification strategy that enhances fluorescent signal intensity at the site of the target antigen.5 In TSA workflows, horseradish peroxidase (HRP), which is commonly linked to a secondary antibody that binds to the primary antibody used to detect the target epitope, catalyzes the localized, covalent deposition of fluorophore-labeled tyramides near the target epitope (Fig. 3). This generates substantially brighter signals than conventional immunofluorescence approaches that rely on directly labeled antibodies.4-5
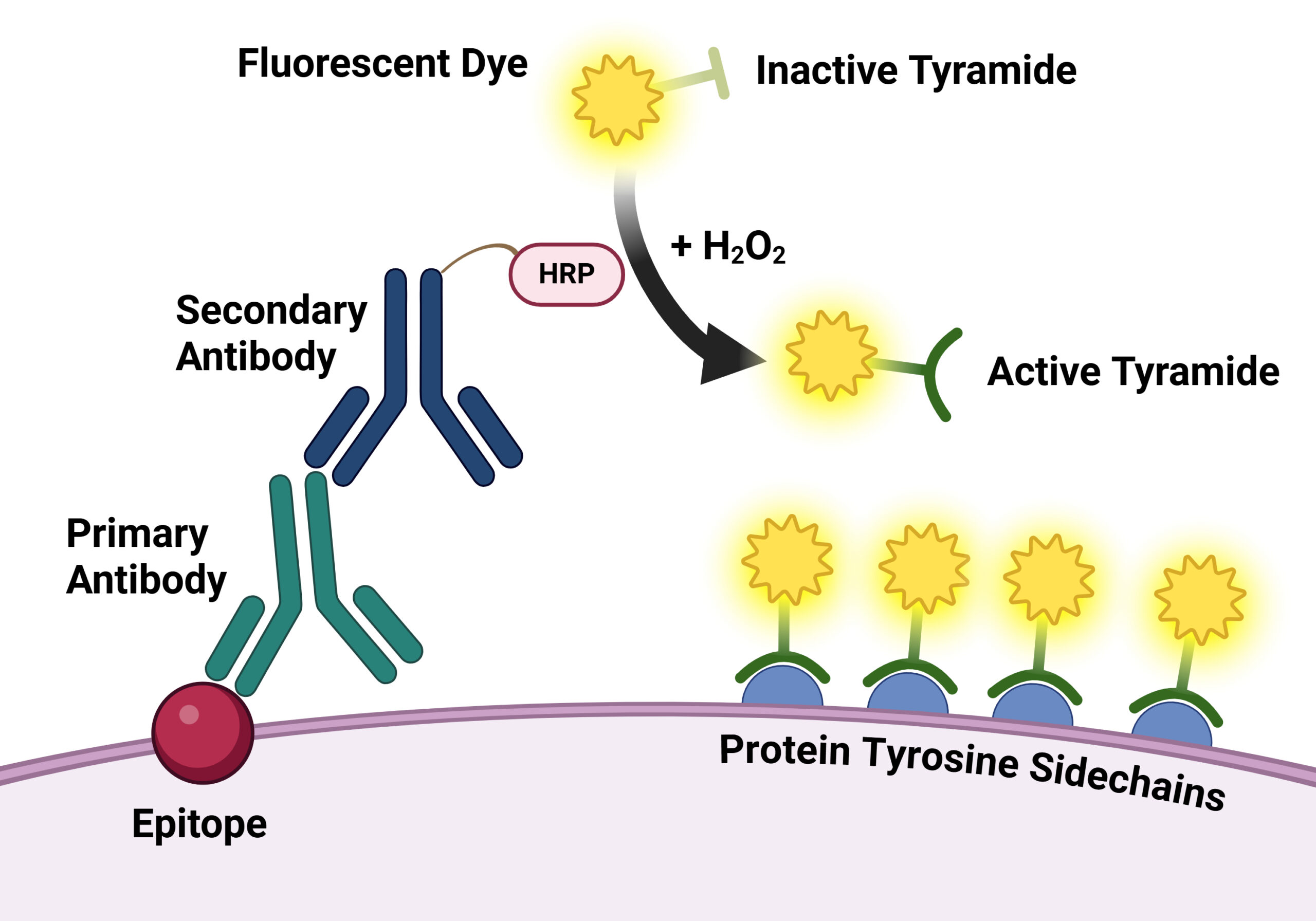
Figure 3. Illustration of the tyramide signal amplification system. Created in BioRender. Davis, J. (2026) https://app.biorender.com/illustrations/697a8eefca57109a76322817.
TSA has become widely used in multiplex immunofluorescence and spatial biology applications because it improves detection sensitivity while supporting high-dimensional imaging workflows.4-5 Enhanced signal amplification can improve visualization of low-expression proteins, phosphorylated targets, and rare cell populations within complex tissues.4-5 For an example of how TSA can be applied in multi-color fluorescence imaging workflows, see this Tech Tip on multi-color fluorescence imaging using TyraMax™ Amplification Kits.
TSA also integrates well with cyclic and iterative imaging approaches. Because deposited tyramide fluorophores become covalently associated with the tissue, antibodies can be stripped or inactivated while the fluorescent signal remains localized within the sample. This enables repeated rounds of staining and imaging on the same tissue section. Although multiple biomarkers can be analyzed simultaneously within each cycle, spectral overlap still limits the total number of fluorophores that can be resolved in a single imaging round. By distributing targets across multiple sequential cycles, cyclic TSA workflows can achieve substantially higher multiplexing capacity than conventional fluorescence imaging alone.4-5
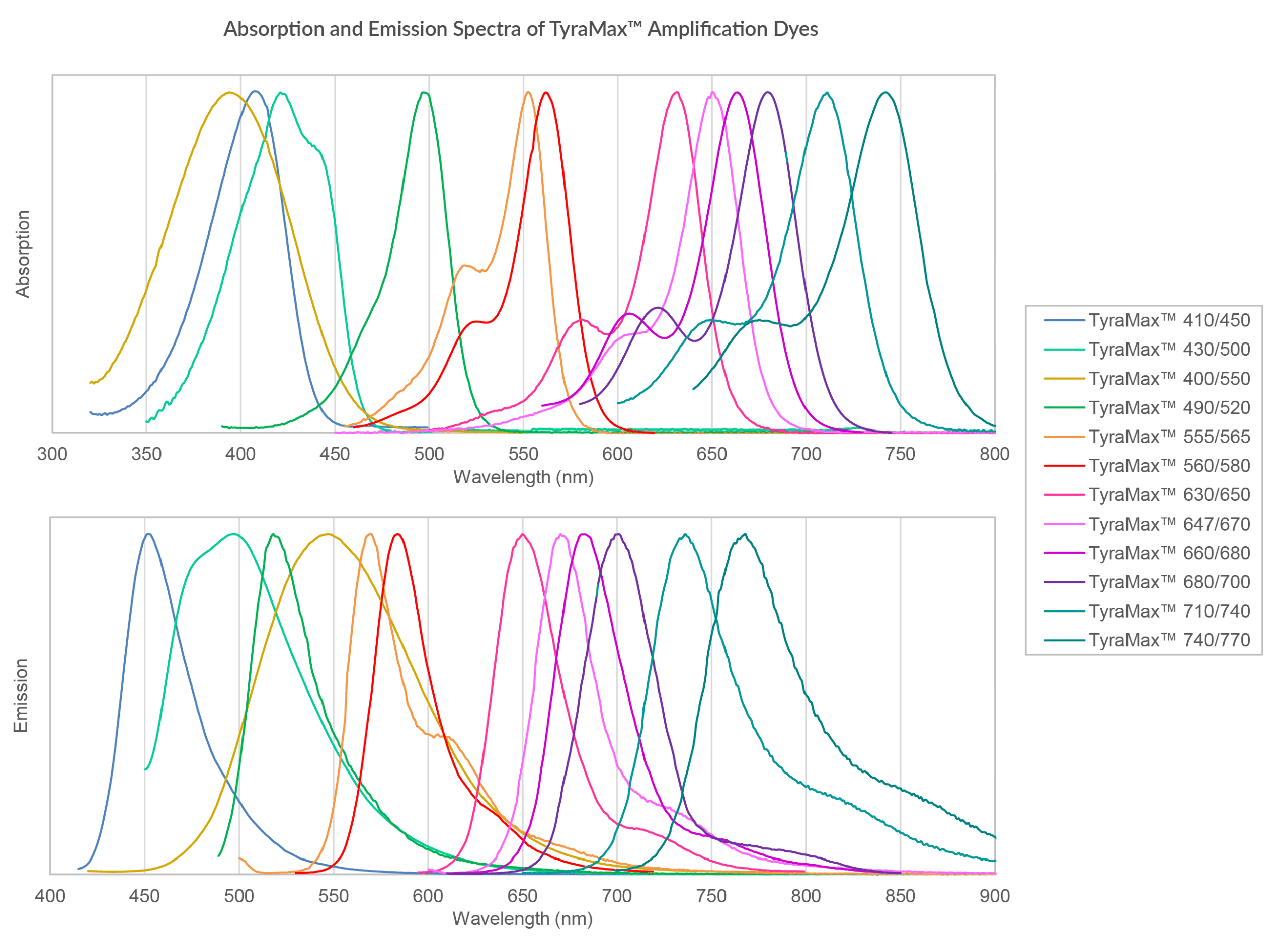
Figure 4. Combined absorption and emission spectra of TyraMax™ Amplification Dyes in PBS.
In practice, the performance of TSA-based workflows is highly dependent on the properties of the tyramide-fluorophore conjugate, as oxidative reaction conditions can impact fluorophore stability. Conventional tyramide reagents may suffer from fluorophore degradation under oxidative conditions or increased background due to nonspecific deposition. TyraMax™ Amplification Dyes and Kits are designed to address these limitations. TyraMax™ Dyes provide enhanced chemostability during HRP-mediated oxidative reactions, helping to preserve signal intensity across multiple staining cycles. In addition, the TyraMax™ portfolio offers one of the broadest selections of tyramide fluorophore options, spanning blue to near-IR wavelengths to support flexible panel design and high-plex imaging workflows (Fig. 4). TyraMax™ Dyes are also engineered for exceptional photostability and brightness, supporting improved signal retention and imaging performance in demanding multiplex spatial biology applications.
As spatial biology workflows become increasingly high-dimensional, the scale and complexity of data generation have created a growing demand for automation and computational analysis tools.6-7 Many multiplex imaging workflows involve repeated cycles of staining, imaging, signal removal, image registration, and data processing, making manual operation labor-intensive and difficult to scale. To address these challenges, researchers are developing increasingly automated spatial biology platforms to improve throughput, reproducibility, and workflow standardization. Automated staining systems, high-content imaging instruments, and integrated analysis pipelines are helping streamline complex spatial omics experiments while reducing user-to-user variability.6-7
At the same time, artificial intelligence (AI) and machine learning approaches are becoming increasingly important for analyzing the large, multidimensional datasets generated by multiplex spatial imaging.6-7 AI-driven image analysis algorithms can assist with:
These computational approaches are particularly valuable in complex tissue environments, where subtle differences in cellular organization, morphology, or biomarker expression may be difficult to identify using conventional analysis methods alone.6 In oncology and translational research, AI-assisted spatial analysis is being explored to improve disease classification, characterize tumor microenvironments, and identify predictive biomarker signatures.7
As automation and AI technologies continue to evolve, they are expected to play an increasingly central role in spatial biology workflows. Together, high-plex imaging, automation, and advanced computational analysis are enabling increasingly comprehensive spatial datasets while improving scalability and analytical precision.6-7
Advances in multiplex imaging, cyclic immunofluorescence, spectral imaging, signal amplification, and AI-driven analysis are transforming how researchers study complex biological systems. By preserving spatial context while enabling increasingly high-dimensional molecular profiling, spatial biology approaches provide deeper insights into cellular organization, signaling networks, and tissue microenvironments than were previously possible. Together, these technologies are driving spatial biology toward increasingly scalable and information-rich workflows that are expanding applications in biomarker discovery, translational research, and precision medicine.
References

Julianne Davis earned an MSc in Behavioral Neuroscience from the University of Washington, Seattle, where she examined the role of memory in cost-based decision-making. She has also studied sensory integration at the Allen Institute and the neural basis of feeding, thirst, and metabolism at the University of California, San Francisco as a research scientist. Currently, she is a Technical Writer and Support Scientist at Biotium.
