 New Products
New Products Earth-Friendly Products
Earth-Friendly Products Biotium Choice Antibodies
Biotium Choice Antibodies Special Offers
Special Offers







Content #1
Content #1
Content #1
Extracellular vesicles (EVs) are known to contain RNA, including microRNA (miRNA) and mRNA, derived from the cell of origin. Studying EV RNA can be useful for many purposes: determining the cellular source of the EVs, for disease diagnostics (Ref. 1), and for identifying oncogenic RNAs (Ref. 2). Since EVs are very small (~30-200 nm in diameter), the amount of RNA cargo that can be isolated and analyzed may be limited. Therefore it is desirable to have robust methods to efficiently purify RNA from the smallest possible number of EVs. Biotium developed the ExoBriteTM EV Total RNA Isolation Kit to isolate RNA of all sizes from EVs with high yield.
In addition to RNA, it has sometimes been reported that EVs also contain DNA. However, most recent publications suggest that the DNA isolated from EVs is external to the EV, and not inside the EV lumen (Ref. 3). During the course of our work developing an RNA isolation kit, we were surprised by the large quantity of DNA that was co-isolated along with the RNA in cell culture-derived EVs, particularly if no DNase treatment was performed. We wondered if this DNA was external to the EVs, and if so, whether it could be removed from the EVs prior to RNA isolation.
To test the hypothesis that the DNA isolated from our EV preparations was external to the EVs (e.g., cell free DNA), we performed a DNase treatment of EVs captured on Biotium’s ExoBriteTM Streptavidin Magnetic Beads coupled to biotinylated tetraspanin antibodies prior to EV lysis for RNA isolation. We compared the yield of recovered RNA to that of EVs that were not treated with DNase, and to samples that underwent the usual on-column DNase treatment after EV lysis. We then quantified the RNA and DNA amounts isolated with these methods using Biotium’s AccuBlue® RNA Quantitation Kit and AccuClear® dsDNA Quantitation Kit, respectively.
We found that treating the EVs with DNase prior to EV lysis removed almost all of the DNA from the EV sample, about as efficiently as the on-column DNase treatment in the usual kit protocol. Interestingly, we found that when the EV DNase treatment was performed prior to the RNA isolation procedure, the RNA yield reproducibly increased. These results suggest that most if not all of the DNA in our EV samples derived from cultured cells is not within the EV, because it can be removed by DNase treatment prior to EV lysis.
EV isolation: EVs were prepared from Jurkat or MCF-7 cells grown for 3 days in serum-free medium. Cell conditioned medium (40 mL) was collected, centrifuged to remove cells and debris, and then the supernatant was concentrated to 2 mL with an Amicon Ultra 10K MWCO ultrafiltration spin column. This concentrated conditioned medium was loaded on an IZON qEV2 size exclusion column and eluted in 8 mL PBS. The isolated EVs were then concentrated to 1 mL with an Amicon Ultra 10K MWCO ultrafiltration spin column. EV concentration was quantified on the ZetaView® QUATT nanoparticle tracking analysis instrument.
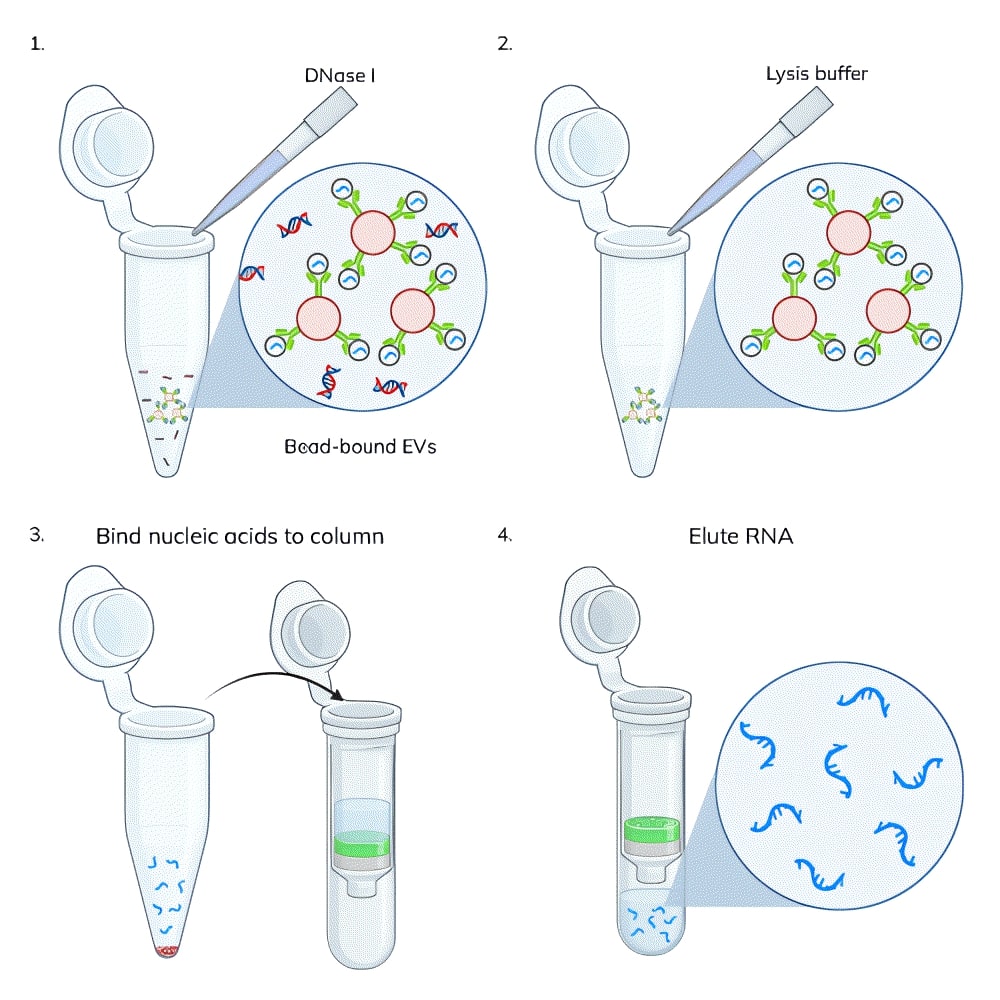
On-bead DNase treatment: EV capture beads were prepared by binding 1 ug of biotinylated tetraspanin antibody (CD81-biotin for Jurkat EVs and CD9-biotin for MCF-7 EVs) to 250 uL of streptavidin beads according to the ExoBriteTM Streptavidin Magnetic Beads protocol. Jurkat cell-derived or MCF-7 cell-derived SEC-purified EVs (3 x 1010 EVs per sample) were added to the capture beads and incubated for 1 hour, rotating. The beads were washed and resuspended in DNase Buffer. The EV/bead samples were each split into two tubes, and DNase I was added to one tube (Figure 1). All of the samples were incubated for 15 minutes, rotating, then washed 3 times with sterile PBS.
Nucleic acid extraction from EVs: Nucleic acids were extracted from 3 x 1010 EVs using the ExoBriteTM EV Total RNA Isolation Kit following the protocol in the product information sheet. Some samples had the typical on-column DNase treatment performed and others did not (Figure 2). Nucleic acids were eluted in 40 uL.
Nucleic acid quantitation: dsDNA was quantified using the AccuClear® Ultra High Sensitivity dsDNA Quantification Kit, using 10 uL of the elution per assay well. ssRNA was quantified using the AccuBlue® Broad Range RNA Quantitation Kit, using 10 uL of the elution per assay well.
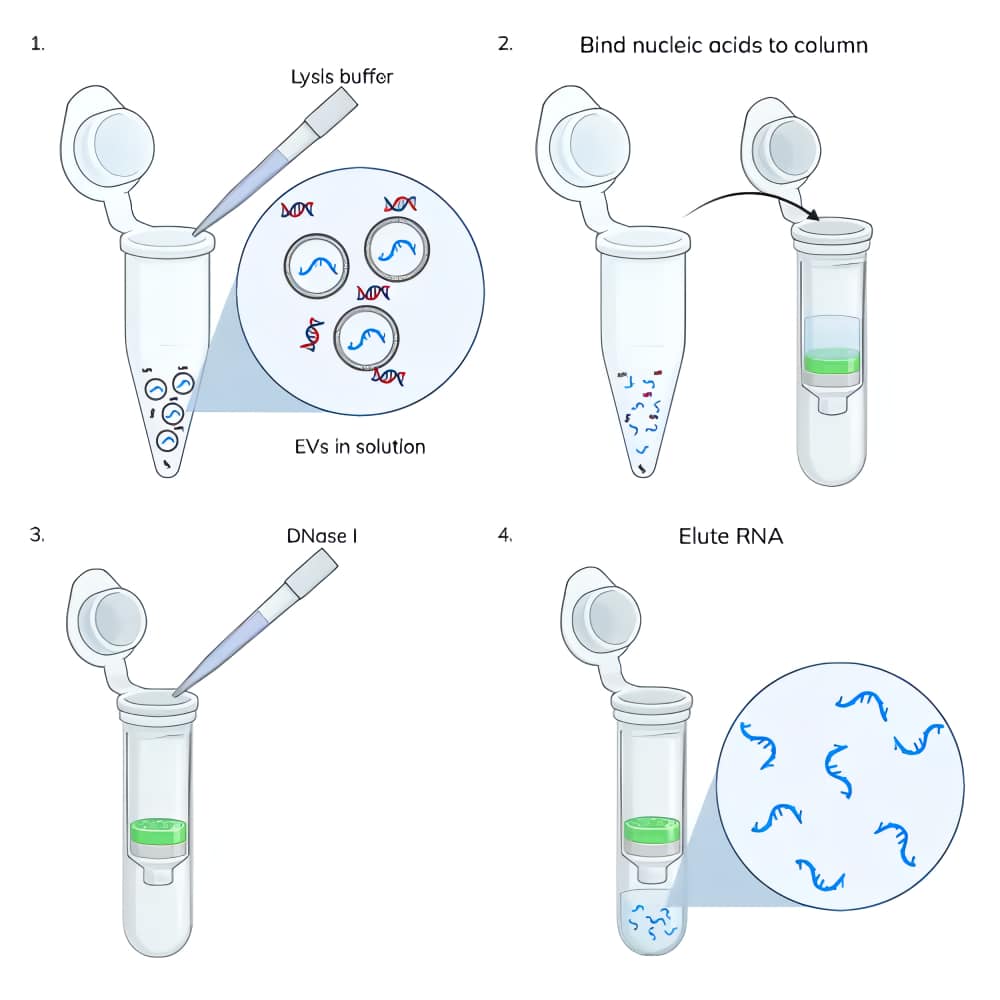
While optimizing the amount of DNase I required to remove DNA during EV RNA extraction, we performed the ExoBriteTM EV Total RNA Isolation Kit protocol with and without on-column DNase I treatment. We were surprised at the relatively large amount of DNA that was co-isolated with RNA (Fig. 3). In two replicate experiments, we found that the amount of RNA isolated from 3 x 1010 EVs was fairly consistent at 40 ng and 50 ng, however the amount of DNA isolated varied more widely at 45 ng and 190 ng. Additionally, the amount of DNA was equal to or greater than the amount of RNA, which we did not expect.
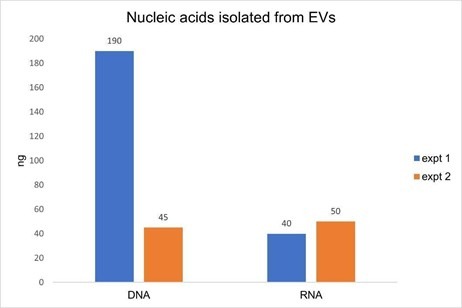
Figure 3. Two replicate experiments were performed in which nucleic acids were isolated from 3 x 1010 Jurkat-derived SEC-purified EVs. In each test, duplicate samples of EVs were processed using the ExoBriteTM EV Total RNA Extraction Kit with the on-column DNase step either omitted or included. DNA was quantified in the no DNase sample using the AccuClear® dsDNA Quantitation Kit, while RNA was quantified in the DNase-treated sample using the AccuBlue® Broad Range RNA Quantitation Kit. The amount of DNA measured was as high or higher than the amount of RNA, in addition to varying widely between experiments, indicative of unexpectedly high levels of DNA in the EV samples.
We hypothesized that the DNA being isolated was external to the EVs (e.g., cell-free DNA) as opposed to being contained within them. To test this, we captured EVs onto magnetic streptavidin beads using biotinylated tetraspanin antibodies (CD81-biotin for Jurkat EVs and CD9-biotin for MCF-7 EVs). Having the EVs captured onto magnetic beads allows the EVs to be treated and washed more easily than purified EVs in solution, because the magnetic beads are easily immobilized on a magnet for buffer exchange and washing. The bead-bound EVs were either untreated or treated with DNase I for 15 minutes using the same buffer and DNase amount as specified for on-column treatment in the extraction kit. The EVs were then washed three times and lysed with the EV Lysis Buffer from the ExoBriteTM EV Total RNA Isolation Kit. The kit protocol was followed for isolation of RNA from the lysate, either with and without the on-column DNase step.
We found that the DNase pre-treatment of bead-bound EVs was as effective as the on-column DNase at removing DNA from an EV-derived nucleic acid sample, removing 90% and 92% of the DNA, respectively (Fig. 4, orange bars). In samples where both bead and column DNase treatments were performed, there was no detectable DNA remaining (there may be some residual DNA below the detection limit of the DNA quantitation assay). We suspect that this additive effect is more likely due to a single DNase treatment not being 100% effective, rather than the residual DNA is inside of the EV lumen, however, we can’t rule that out without further testing.
In examining this data, we noticed an interesting phenomenon: in both of the samples where the EVs were treated with external DNase prior to lysis (i.e., “Bead DNase” and “Col+bead DNase”) the yield of isolated RNA was higher than the no-DNase sample, by 28-38% (Fig. 2, blue bars). This experiment was performed twice with EVs isolated from two different cell lines (Jurkat and MCF-7), and the yield improvement with pre-DNase treatment was observed in both instances.
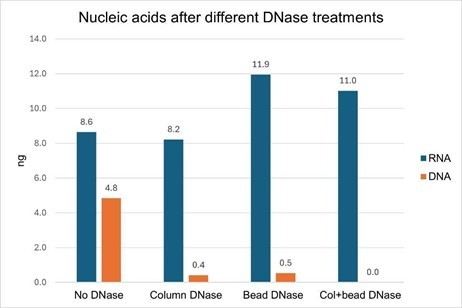
Figure 4. Jurkat-derived SEC-purified EVs (3 x 1010) were captured onto magnetic streptavidin beads with a CD81-biotin antibody. Two of the EV samples (“Bead DNase” and “Col+bead DNase”) were treated with DNase I for 15 minutes at room temperature and two samples (“No DNase” and “Column DNase”) were untreated. All of the samples were washed 3 times and then lysed. Nucleic acids were isolated with or without on-column DNase I treatment. Both DNase treatments (on beads and on column) removed nearly all of the DNA from the isolated nucleic acid, and no DNA was detectable when both DNase treatments were performed. When bead-bound EVs were treated with DNase, the RNA yield was increased.
This is not the first report indicating that most or all DNA in an EV sample is external to the vesicles (Ref. 3), and our data adds to that mounting evidence. While we did see an additive reduction in DNA when both DNase treatments were performed, we did not perform controls where two on-bead or two on-column treatments were done. Nevertheless, it is likely that a single DNase treatment is not sufficient to remove all of the DNA in a sample, and two sequential DNase treatments appear to be more effective; we have seen this to be true in other applications (data not shown). More testing would be needed to clarify this.
DNase-treating the surface of EVs and thus removing most of the DNA in the sample prior to EV lysis resulted in an increase in the RNA yield from those EV samples (Figure 1). We hypothesize that there is competitive binding to the spin column between DNA and RNA in the lysate. It is known that DNA binds to silica membranes with higher affinity than RNA because it is more highly charged so this competition is not unexpected.
These results suggest that one way to generate high yield and high purity RNA from EVs is to perform a DNase treatment of the EVs prior to lysis and nucleic acid purification. However, there are some caveats. This procedure is currently only practical for EVs that are captured onto beads or in some other manner that allows washing, and since a capture molecule such as an antibody may not capture every EV in a sample, some EVs will likely be lost, and the remaining EV population may display a bias based on the capture molecule. However, as more efficient EV capture technologies are developed, this process will become more practical for broad use.

Content #1
Content #1
Content #1
Content #2
Content #3