 New Products
New Products Earth-Friendly Products
Earth-Friendly Products Biotium Choice Antibodies
Biotium Choice Antibodies Special Offers
Special Offers



 Powered by Bioz
Powered by Bioz








Content #1
Content #1
Content #1
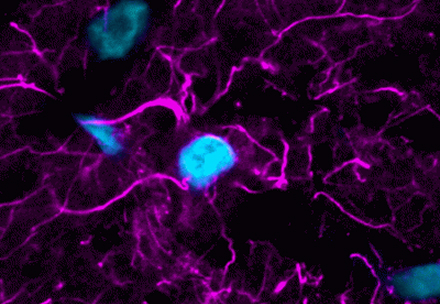
Cell-permeant probes for imaging the microtubule cytoskeleton in live cells. Available with blue, green, or far-red fluorescence.
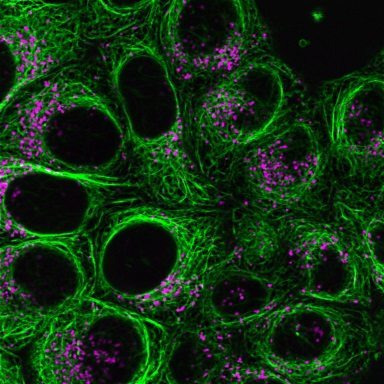
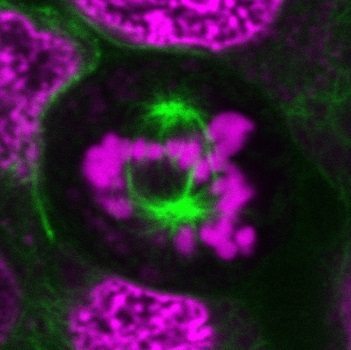
ViaFluor® Live Cell Microtubule Stains are cell-permeant probes for imaging the microtubule cytoskeleton in live cells. They are simple, rapid and sensitive stains that can be imaged without a wash step.
ViaFluor® Live Cell Microtubule Stains bind to polymerized tubulin and may be used to visualize microtubules in live cells for up to 24 hours in immortalized cell types. However, the probes may show cytotoxicity and cell cycle arrest with longer incubation times. The stains are supplied as a 1000X stock solutions in DMSO, and include a vial of 100 mM VRP, an efflux pump inhibitor that may improve probe retention and staining in some cell types.
ViaFluor® Live Cell Microtubule Stains cannot be fixed after staining, and cannot be used with fixed cells or tissues. The probes are not predicted to bind bacterial or yeast microtubules.
Learn more about our other Fluorescent Cellular Stains for nucleus, mitochondria, lysosomes, or see our selection of Microbiology Stains & Kits.
| Dye | Ex/Em | Catalog no. | Size (1000X in DMSO)* | Features |
|---|---|---|---|---|
| ViaFluor® 405 | 408/452 nm | 70064-T | 10 uL | Blue fluorescence for the DAPI channel |
| 70064 | 50 uL | |||
| ViaFluor® 488 | 500/515 nm | 70062-T | 10 uL | Green fluorescence for the FITC channel |
| 70062 | 50 uL | |||
| ViaFluor® 647 | 650/675 nm | 70063-T | 10 uL | Far-red fluorescence for the Cy®5 channel Compatible with SIM or STED |
| 70063 | 50 uL |
The mechanism of binding for RedDot™ 1 and RedDot™ 2 to DNA has not been characterized. However, based on the dye structure, it may bind by a similar mechanism as DRAQ®5, which has been reported in the literature to be a concentration-dependent intercalator and minor groove binder.
DRAQ is a registered trademark of Biostatus, Ltd.
Mitochondrial dyes, including MitoView™ Mitochondrial Dyes, are positively charged and lipophilic. They passively diffuse across cellular membranes and are presumed to accumulate in the mitochondrial matrix due to the proton gradient in the mitochondria (for a detailed review, see Cytometry Part A79A: 405-425, 2011).
However, some dyes are still retained in mitochondria after depolarization. Our dye chemists hypothesize that this is because some of the dyes are more lipophilic than others. Once they accumulate in the mitochondria because of their charge, they are less likely to diffuse back into the cytoplasm due to their hydrophobicity, even after the proton gradient that attracted them is dissipated by mitochondrial depolarization. Probably they associate with the mitochondrial membranes instead.
The so-called potential-independent dyes like MitoView™ Green, MitoTracker® Green, and Nonyl Acridine Orange are much more hydrophobic than potential-responsive dyes like MitoView™ 633, Rhodamine 123, and JC-1. The former dyes are retained after mitochondrial depolarization, and can be used to measure mitochondrial mass independent of potential. However, it would be more accurate to call these dyes relatively potential-insensitive, rather than potential-independent, because mitochondrial potential still plays a role in their localization. These dyes have been reported to show some loss of signal upon depolarization (Cytometry 39(3):203-10, 2000).
There is another class of mitochondrial dyes that accumulate in mitochondria based on charge, but also have a reactive group that can covalently link the dye to protein targets within the mitochondria, allowing them to be well-retained after fixation and permeabilization. Our MitoView™ Fix 640 is this type of dye.
Some dyes, like MitoView™ Green can stain mitochondria in cells that are already fixed. The mechanism by which this occurs is not well-understood. After fixation, there should be no proton gradient in the mitochondria to attract the dyes at all. Our chemists suspect that there may be some residual membrane potential in fixed mitochondria that is not due to the proton gradient (which would disappear following fixation), but instead arises from uneven distribution of proteins that have different isoelectric points (net charge). There are reports that the net charge of resident proteins in organelles differs based on the pH of the cellular compartment (Proc Natl Acad Sci USA 115(46):11778-11783, 2018). Charge differences may be sufficient to attract cationic lipophilic dyes to mitochondria in the absence of a proton gradient, due to a combination of weak electrostatic and hydrophobic interactions with mitochondrial proteins and membranes.
However, currently there is no direct evidence to suggest this is the mechanism for MitoView™ Green staining of fixed cells. There may be other targets that the dye is binding. For example, Nonyl Acridine Orange is reported to bind cardiolipin, a lipid that is enriched in mitochondrial membranes. It's possible that MitoView™ Green binds to particular molecules in mitochondria with some degree of specificity. However, staining of fixed cells with mitochondrial dyes generally is not as specific as staining of live cells. That's why we recommend using mitochondrial marker antibodies instead of dyes to stain fixed cells when possible.
Mounting medium can alter the staining of lipophilic dyes like LipidSpot™, due to interaction of the dyes with glycerol or other components that help form the interface between the coverslip and slide. The antifade compounds in mounting medium are generally compatible with the dyes. In our tests, LipidSpot™ staining was well preserved in EverBrite™ Mounting Medium (catalog. nos. 23001/23002) for up to 24 hours after mounting, but lipid droplet size and staining intensity were somewhat altered after samples were stored in mounting medium for several days. Therefore, if mounting medium is required to image samples, we'd recommend imaging as soon as possible after mounting.
LipidSpot™ is not compatible with FluoroShield mounting medium (staining is lost immediately after mounting). We have not tested other types of mounting medium.
